Contents
Net Community Productivity: in situ O2/Ar Measurements
In the surface ocean, net community productivity (NCP) is a function of photosynthesis and community respiration according to the following approximate stoichiometry,
(1) 106 CO2 + 16 HNO3 + H3PO4 + 122 H2O ↔ (CH2O)106(NH3)16H3PO4 + 138 O2
NCP can be estimated from the net change in the inventories of several upper ocean properties, notably of oxygen, organic and inorganic carbon, nitrogen and phosphorus. In equation (1), if the forward reaction (photosynthesis) exceeds the reverse reaction (respiration), the system is net autotrophic, which leads to an increase in oxygen and a decrease in inorganic carbon within the mixed layer. This net autotrophy is characterized by a net accumulation and sinking of organic carbon, in the form of particulate and dissolved organic carbon. On the other hand, if heterotrophy exceeds autotrophy within the mixed layer, oxygen decreases and inorganic carbon increases. Because oxygen and inorganic carbon have a gaseous component, a gas exchange flux component must be included in their mass balance budgets.

Oceanic carbon and nutrient cycling
NCP from oxygen mass balance
Because of the central role O2 plays in biological redox reactions in the surface ocean, the biological oxygen budget first and foremost reflects the net energetic balance. An ecosystem where the photochemical energy input is greater than the loss of potential energy from organic matter oxidation has a positive internal energy change. The oxygen balance also mirrors net organic carbon production because of the stoichiometric coupling of oxygen and carbon.
Oxygen in the mixed layer is however also influenced by physical processes such as bubble injection, temperature and pressure changes. Because argon (Ar) has similar solubility properties as oxygen, the oxygen derived from physical processes can be estimated from the argon concentration relative to its saturation ([Ar]sat). The oxygen derived from biology ([O2]b) is equal to the total oxygen minus the oxygen derived from physical processes ([O2]p):
(2) [O2]b = [O2] – [O2]p ≈ [O2] – ([Ar]/[Ar]sat) = ([Ar]/[Ar]sat) [O2]sat Δ(O2/Ar)
where
(3) Δ(O2/Ar) = [(O2/Ar)/(O2/Ar)sat-1]
and is defined as the biological oxygen supersaturation. Analogous to the apparent O2 utilization (AOU), [O2]b is a synthetic tracer of biological activity within the mixed-layer, and can be negative if the mixed-layer is net heterotrophic.
Neglecting vertical mixing, the biological oxygen net flux within the mixed-layer of the ocean is a function of the mass balance between net community oxygen production and exchange of biological oxygen with the atmosphere,
(4) MLD d[O2]b/dt = NCP – kO2[O2]b
where MLD is the mixed-layer depth, and kO2 is the gas exchange coefficient for oxygen (aka piston velocity). Substituting [O2]b in Eq. (4) with the term on the right-hand side of the equality sign in Eq. (2), NCP under steady-state conditions can be approximated by
(5) NCP ≈ kO2 [O2]sat Δ(O2/Ar) ([Ar]/[Ar]sat) ≈ kO2 [O2]sat Δ(O2/Ar)
We ignore the argon saturation in Eq. (5) because it is close to one; it does not significantly alter the NCP estimates and is within the measurement errors. Under net auto (hetero)-trophic mixed-layer conditions, the net oxygen flux is out (in) of the ocean.

NCP observations based on O2/Ar measurements in the world’s oceans
Based on Eq. (5), NCP can be derived from 1) the oxygen concentration at saturation ([O2]sat), 2) a gas exchange coefficient (kO2), and 3) the biological oxygen supersaturation (Δ(O2/Ar)). We calculate the oxygen solubility from atmospheric pressure, temperature, and salinity measurements. The piston velocity for oxygen is estimated based on a windspeed reanalysis or satellite dataproducts and a windspeed parameterization. The piston velocity is weighted for the windspeed history at the collection site. Finally, the oxygen biological supersaturation is measured continuously by Equilibrator Inlet Mass Spectrometry (EIMS), a method previously described by Cassar et al. (2009). Briefly, seawater from the ships underway system is pumped through a gas equilibrator, the headspace of which is connected to a quadrupole mass spectrometer for continuous elemental O2/Ar ratio measurements, from which the biological oxygen supersaturation is estimated. From the O2/Ar supersaturation, the piston velocity, and the oxygen concentration at saturation, the net biological oxygen flux across the ocean surface is estimated based on Eq.(5).
With collaborators, we measure NCP in the world’s ocean to 1) constrain the magnitude, 2) assess the variability and 3) investigate the controls on productivity and carbon cycling in the surface ocean. See some of our publications for more information.
Satellite estimates of NCP
We derived two statistical algorithms for predicting global oceanic NCP from satellite observations. To calibrate these two algorithms, we compiled a large dataset of in situ O2/Ar-NCP and remotely-sensed observations, including Sea Surface Temperature (SST), Net Primary Production (NPP), phytoplankton size composition, and inherent optical properties. The first algorithm is based on genetic programming (GP) which simultaneously searches for the optimal form and coefficients of NCP equations. We find that several GP solutions are consistent with NPP and SST being strong predictors of NCP. The second algorithm uses support vector regression (SVR) to optimize a numerical relationship between O2/Ar-NCP measurements and satellite observations. Both statistical algorithms can predict NCP relatively well, with a coefficient of determination (R2) of 0.68 for GP and 0.72 for SVR, which is comparable to other algorithms in the literature. However, our new algorithms predict more spatially uniform annual NCP distribution for the world’s oceans and higher annual NCP values in the Southern Ocean and the five oligotrophic gyres. We have used also used other machine learning methods (artificial neural networks; self-organizing maps; Bayesian hierarchical model) to derive algorithms at basin scales, and applied these products to understand controls on NCP at global, basin, and regional scales (e.g. Southern Ocean & Western Antarctic Peninsula). See some of our publications for more information.
Gross Primary Productivity: Dissolved Oxygen Triple Isotopes
We estimate gross primary productivity (GPP) in the oceanic mixed-layer based on the triple isotope composition of dissolved O2. Oxygen in the surface ocean has two main sources: the atmosphere and the photosynthetic splitting of water. These sources have distinct isotopic signatures. Atmospheric oxygen has a characteristic isotopic signature derived from a stratospheric reaction with a mass independent fractionation between O2, CO2, and ozone. Mass-independent fractionation is manifested in a non-zero 17Δ:
(6) 17Δ = ln(0.001 δ17O + 1) – 0.516ln(0.001 δ18O+1) ~ δ17O – 0.516 δ18O
The overall isotopic signature of oxygen in the surface ocean thus depends on the weighted abundance of atmospheric and photosynthetic oxygen. The sinks for dissolved O2 in the surface ocean are respiration and gas exchange. These processes fractionate O2 mass dependently and therefore do not change 17Δ. GPP is the rate of photosynthetic O2 production required to maintain the observed departure of 17Δ from atmospheric equilibrium against dissipation from gas exchange.
Atmospheric O2/N2 and Ar/N2 measurements

Atmospheric O2/N2 and Ar/N2 stations
Atmospheric O2/N2
Measurements of atmospheric O2/N2 ratios provide a unique insight into the sinks of fossil fuel CO2, marine carbon cycling, productivity fluxes, and ocean circulation. For example, the seasonal cycle in “atmospheric potential oxygen”, a synthetic tracer combining the atmospheric O2 and CO2 signals and which is for all intent and purposes conservative with respect to the land biosphere, reflects the balance between marine net community productivity and ventilation of deep waters.

Atmospheric O2/N2 measurements. CO2 measurements performed by NOAA lab (Boulder)
Concurrent measurements of atmospheric O2 and CO2 can also be used to assess the partitioning of anthropogenic CO2 uptake between the land biosphere and the oceans. The observed increase in atmospheric CO2 concentration is less than predicted based on anthropogenic fossil fuel emissions. The difference between the predicted and the observed CO2 concentration represents a global CO2 sink, attributed to increases in land biosphere biomass and oceanic CO2 solubility. The atmospheric CO2 signal is obviously affected by both of these processes and by fossil fuel burning. On the other hand, the atmospheric oxygen signal only represents changes in the land biosphere biomass and fossil fuel burning.
Atmospheric Ar/N2

Comparison of atmospheric Ar/N2 observations and model predictions
The atmospheric ratio of argon to nitrogen (Ar/N2) varies seasonally because, in part, solubilities of these gases change with temperature. Since Ar is approximately twice as soluble as N2, the atmospheric Ar/N2 ratio increases as the surface oceans warm (i.e., spring, summer) and degas. The opposite occurs during cooling (i.e., fall, winter). At a given atmospheric sampling site, the Ar/N2 seasonal amplitude is a function of the oceanic temperature change, the air-sea gas exchange rate, and the dilution of the Ar/N2 signal by atmospheric transport and mixing. In this respect, atmospheric Ar/N2 is a valuable tracer of the complex interplay of upper ocean heating, air-sea gas flux kinetics, and atmospheric transport dynamics. Temporal (e.g., interannual) variability in these processes should be reflected in the atmospheric Ar/N2 record, and differences between sampling sites may highlight spatial variations in these processes. Furthermore, the rise in sea surface ocean temperature associated with global warming should manifest itself as a secular increase in the Ar/N2 ratio in air. The latter is difficult to observe because of the large noise to signal ratio. Comparing Ar/N2 observations with simulations from paired ocean-atmosphere models tests the robustness of simulations of upper ocean mixing, air-sea heat fluxes and atmospheric transport.
Contact Nicolas Cassar (nicolas.cassar_at_duke.edu) or Michael Bender (bender_at_princeton.edu) for atmospheric O2/N2 or Ar/N2 data use.
Algal Ecophysiology
We investigate several aspects of algal physiology with implications for the biogeochemical cycle of carbon in the ocean, global climate change, and paleoceanography. Some of our current research interests include carbon-concentrating mechanisms (CCMs) and β-carboxylation in marine phytoplankton and their potential influence on photosynthetic carbon isotope fractionation, iron limitation, and diatom morphometrics.
Carbon Assimilation in Marine Phytoplankton
Important but controversial laboratory and field studies suggest that marine primary productivity could be enhanced in response to increasing atmospheric CO2. Although inorganic carbon is abundant in seawater, most of it is in the form of bicarbonate (HCO3–) and carbonate (CO32-), which are not readily available to marine photoautotrophs. Ribulose 1,5-bisphosphate carboxylase-oxygenase (RubisCO), the principal enzyme that fixes inorganic carbon during photosynthesis, is less than half saturated at normal seawater CO2 concentration. Hence, if phytoplankton species are unable to access inorganic carbon pools other than CO2, they could in fact be CO2 limited. Many phytoplankton species have evolved carbon-concentrating mechanisms to compensate for RubisCO’s catalytic inefficiency. Active uptake of HCO3– and CO2 through the cell membrane, active transport of inorganic carbon from the cellular membrane to the chloroplasts, and active, conversion of HCO3– to CO2 by a carbonic anhydrase-like mechanism are putative CCMs. We can gain insight into these various pathwayswith cellular energy minimization models, enzymatic assays, stable and radio-isotope carbon isotope fractionation and disequilibrium experiments. Most of our experiments are performed on chemostats under various CO2 concentrations. In a nutrient limited chemostat, the phytoplankton growth rate at steady state is equal to the dilution rate of the growth chamber.The partial pressure of CO2 in the gas used to aerate the growth chamber is controlled using mass-flow controllers to adjust the flow rates of tank CO2 and CO2-free medical air.
Carbon Isotopes in Marine Phytoplankton

Chemostat (continuous-growth cultures) experiments
During the photosynthetic process, marine phytoplankton preferentially fix 12C relative to 13C. The extent of fractionation between 12C and 13C is dependent on CO2 availability. As a result, the isotopic signature of phototrophic organic matter in marine sediments, such as δ13C37:2 of alkenones produced by some coccolithophores, can potentially be used to reconstruct atmospheric CO2 concentration over geological timescales. Unfortunately, a multitude of confounding factors may influence the dependence of photosynthetic carbon isotope fractionation (εp) on CO2. Variations in the degree of dependence of fractionation on CO2 call for more laboratory work. Few studies have looked at carbon-concentrating mechanisms (CCMs) in haptophytes and how these mechanisms affect εp. The contribution of inducible CCMs to εp in E. huxleyi can be further investigated by performing radiocarbon isotope disequilibrium experiments in conjunction with stable carbon isotope fractionation measurements on continuous growth cultures of E. huxleyi. A quantitative understanding of the still rather elusive physiological parameters that modulate εp is mandatory before the sedimentary carbon isotope record can be confidently interpreted for paleo-CO2 reconstruction. See some of our publications for more information.
Method Development
Scientific questions such as the ones discussed above have led us to develop innovative methods, described below.
Equilibrator Inlet Mass Spectrometry (EIMS)

Equilibrator Inlet Mass Spectrometry

Inlet of EIMS
We designed a new method for continuous shipboard measurements of O2/Ar by equilibrator inlet mass spectrometry (EIMS). From these measurements and an appropriate gas exchange parameterization, NCP can be estimated at unprecedented spatial and temporal resolution. This new instrument has now been deployed successfully several times in the Southern Ocean, Bering Sea, and the Pacific and Atlantic Ocean.
In the EIMS configuration, seawater from the ship’s continuous intake flows through a cartridge enclosing a gas-permeable microporous membrane contactor. Gases in the headspace of the cartridge equilibrate with dissolved gases in the flowing seawater. A fused-silica capillary continuously samples headspace gases, and the O2/Ar ratio is measured by mass spectrometry. The ion current measurements on the mass spectrometer reflect the partial pressures of dissolved gases in the water flowing through the equilibrator. Calibration of the O2/Ar ion current ratio (32/40) is performed automatically every 2 h by sampling ambient air through a second capillary. N2/Ar and pCO2 may potentially also be measured by EIMS.

EIMS nXT
EIMS. The large seawater reservoir (A) sits in a sink. After going through an inline coarse filter (500 μm pore size), seawater flows into the inner reservoir (B) at a rate of 3-5 L min-1 (large arrow). Most of the water running into B overflows into A, which is for thermoregulation of the system. A small fraction (100 mL min-1) of the high flow rate is pulled with a gear pump through a filter sleeve (C), with 100 and 5 μm pore size on the outside and inside, respectively. From the gear pump, the seawater flows through the equilibrator (D). The equilibrator sits in reservoir A to keep its temperature identical to that of the incoming seawater. A capillary, attached to the headspace of the equilibrator, leads to a multiport Valco valve. This valve alternates between admitting gas from the equilibrator and ambient air to the quadrupole mass spectrometer. An optode (not shown) in container B measures total oxygen saturation. Also not shown is a water flow meter located downstream of the equilibrator and thermocouples monitoring temperatures throughout the system.
Dual Isotope Cavity Ring-Down Laser Absorption Spectroscopy

Underway DIC method
In collaboration with Kuan Huang (Picarro), Michael Bender (Princeton University) and other collaborators, we have developed a new method for continuous measurement of dissolved inorganic carbon (DIC). The method is based on dual isotope dilution and cavity ring-down spectroscopy (DID-CRDS). In this method, seawater is continuously sampled and mixed with a flow of NaH13CO3 solution that is also enriched in deuterated water (the spike). The isotopic composition of CO2 (δ13Cspiked_sample) derived from the DIC in the mixture, and the D/H ratio of the mixed water (δDspiked_sample), are measured by CRDS analyzers. See Huang et al. (2015) for more information.
ARACAS: N2 fixation in terrestrial environments

ARACAS
The most common currency for estimating N2 fixation is acetylene reduction to ethylene. Real-time estimates of nitrogen fixation are needed to close the global nitrogen budget and remain a critical gap in both laboratory and field experiments. We present a new method for continuous real-time measurements of ethylene production: Acetylene Reduction Assays by Cavity ring down laser Absorption Spectroscopy (ARACAS). In ARACAS, air in the headspace of an incubation chamber is circulated with a diaphragm pump through a cavity ring-down ethylene spectrometer and back to the incubation chamber. The new approach offers several benefits compared to the conventional detection of ethylene by flame ionization detector gas chromatography. First, the detection of acetylene reduction to ethylene is non-intrusive and chemically non-destructive, allowing for real-time measurements of nitrogenase activity. Second, the measurements are made instantaneously and continuously at ppb levels, allowing for observation of real-time kinetics on time intervals as short as a few seconds. Third, the instrument can be automated for long time periods of measurement.
We illustrate its use for measuring N2 fixation using two species, the diazotrophic bacterium Azotobacter vinelandii and the lichen Peltigera praetextata. Using ARACAS, we also characterize short-term kinetics of N2 fixation by cyanobacteria on moss carpets from warm and cold temperate forests. We report the identification of a heretofore unknown multispecies true-moss-cyanobacteria diazotrophic association. We demonstrate that short-term change in abiotic variables greatly influences N2fixation. We also show that difference in relative proportion of two epiphytic diazotrophs is consistent with divergent influences of temperature on their N2 fixation kinetics. See some of our publications for more information.
FARACAS: N2 fixation in freshwater and oceanic environments
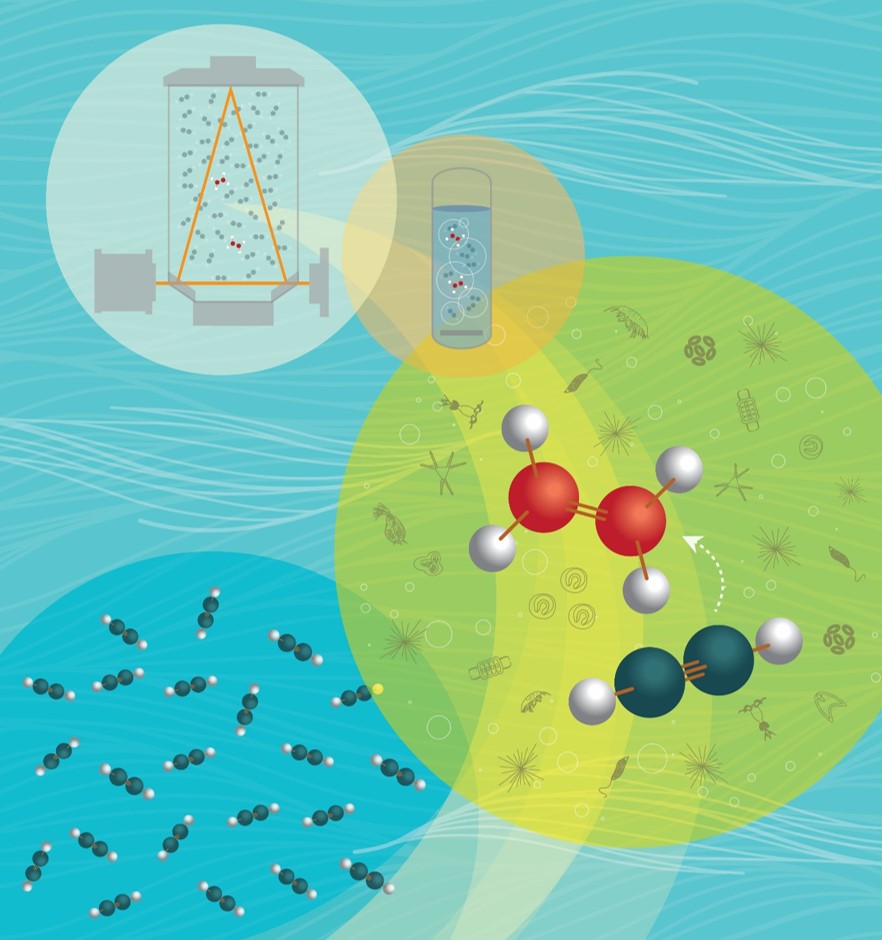
FARACAS Concept
Because of the difficulty in resolving the large variability of N2 fixation with current methods which rely on discrete sampling, the development of new methods for high resolution measurements is highly desirable. We present a new method for high-frequency measurements of aquatic N2 fixation by continuous flow-through incubations and spectral monitoring of the acetylene (C2H2, a substrate analog for N2) reduction to ethylene (C2H4). In this method, named “Flow-through incubation Acetylene Reduction Assays by Cavity ring-down laser Absorption Spectroscopy” (FARACAS), dissolved C2H2 is continuously admixed with seawater upstream of a continuous-flow stirred-tank reactor (CFSR) in which C2H2 reduction takes place. Downstream of the flow-through incubator, the C2H4 gas is stripped using a bubble column contactor and circulated with a diaphragm pump into a wavelength-scanned Cavity Ring Down laser absorption Spectrometer (CRDS). Our method provides high-resolution and precise mapping of aquatic N2 fixation, its diel cycle, and its response to environmental gradients, and can be adapted to measure other biological processes. The short-duration of the flow-through incubations without preconcentration of cells minimizes potential artefacts such as bottle containment effects while providing near real-time estimates for adaptive sampling. We expect that our new method will improve the characterization of the biogeography and kinetics of aquatic N2 fixation rates. See some of our publications for more information.

FARACAS process flow and mind map diagrams

You must be logged in to post a comment.